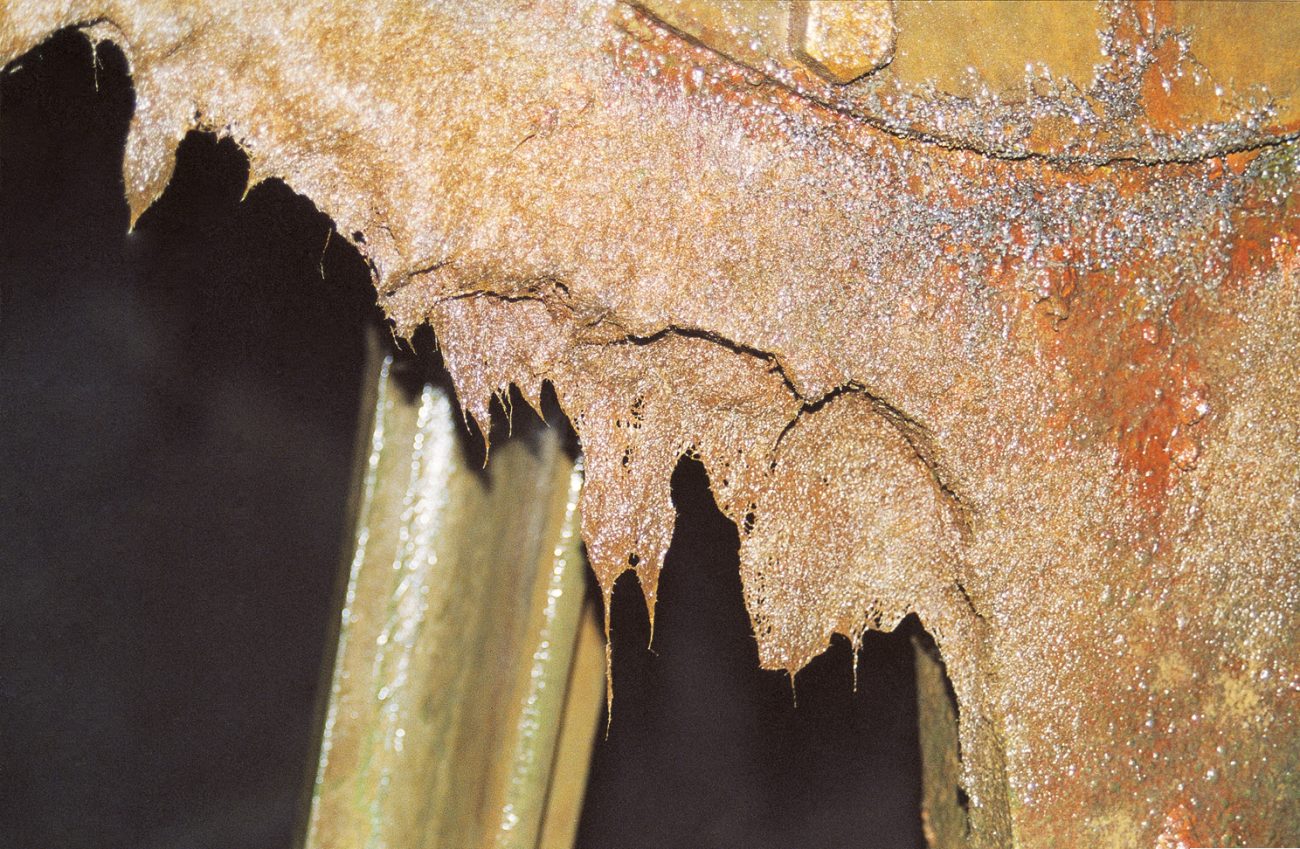
Living lace
Crisp and wafer-thin, a colony of the lace coral Hippellozoon novaezelandiae looks good enough to eat. But like all bryozoans it produces toxic compounds to deter other creatures from nibbling it or settling on top of it. Such chemical defences are one of the reasons for the success of bryozoans—for these animals are among the commonest marine invertebrates. With nearly 1000 species, New Zealand possesses one of the richest bryozoan diversities in the world.
Thank you, Mount Albert Grammar School. That’s where it all started—in the fifth form, with a collection of preserved animals and Ralph Buchsbaum’s two-volume paperback classic, Animals Without Backbones. The pages in my copy have yellowed with the years, but the text is still as fascinating to me now I’m a professional zoologist as it was in my pimply youth. And the photographs! Regenerating hydras and cross-eyed planarian worms, comb jellies, human legs bloated by elephantiasis, leeches and liver flukes, king crabs and chitons, rotifers and . . . bryozoans. There weren’t any bryozoans in the class collection. I do remember comb jellies, though, which disintegrated when I whirled them around in the jar. (Yes, sorry, that was me.)
Our family lived in New Lynn, in Auckland’s western suburbs. The property had a river running through it. A creek, actually (except when in spate), but it had neat creatures in it, such as planarian worms, leeches, shrimps, orange nemertean worms and bryozoans just like in the books.
When magnified, creek life became a magical world to me: an undiscovered realm, a cosmos of possibilities, an art show of shapes, colours, patterns and movements. I don’t know what it is about small invertebrates that captivates some people, but from the time I first watched live bryozoans under a low-power microscope I was hooked.

I’m not the only one. In 1856, English naturalist George Allman expressed his own admiration:
Among the most beautiful and interesting forms of invertebrate animals are those strange phytoidal productions . . . Bryozoa. They are chiefly inhabitants of the sea, where they may be witnessed under numerous plantlike guises; now spreading like a lichen over submerged stones, or old shells, or the broad fronds of Laminaria and other sea-weeds; now forming soft, irregular, fungus-like masses, or hard, calcareous branchy growths, like diminutive trees; and now again presenting the appearance of the most delicate and exquisitely formed sea-weed or moss, offering, even to the unassisted eye, in the endless repetition of the same form, objects of surpassing symmetry and beauty.
They are protean things, bryozoans. Witness their varied common names. The bushy ones used to be called “moss animals” (which is the literal meaning of “bryozoa,” from two Greek words) and the flat encrusting ones “sea mats.” I prefer the term “lace corals,” for some indeed look like fragile lace creations, and even the encrusters form pretty sheets over seaweeds, stones and shells. But no one common name is adequate for all, so the scientific name Bryozoa (“bryozoan” in more informal usage) is probably the best all-round term. And that’s a pity in a way, too, because the now disused name Polyzoa (“many animals”) is more usefully descriptive: all bryozoans, with the exception of one or two genera that may consist of a single individual for part or most of their lives, are colonies of a few to millions of individuals. In today’s parlance we would call them “modular” or “clonal” organisms.
Technical and common names aside, who has actually heard of bryozoans, apart from marine biologists? I think they’re the world’s best-kept secret. I used to say to my children, “If anyone asks what your father does, tell them he studies things that grow on the bottom of boats.”
There’s more to it than that, of course, but it so happens that bryozoans are among the world’s most troublesome marine-fouling organisms. The larvae of some bryozoan species are highly tolerant of the heavy metals contained in antifouling coatings that inhibit the settlement and growth of other animals.

Both bushy and flat bryozoans are a problem. Moreover, other organisms, such as barnacles, which may be less tolerant of antifouling chemicals than bryozoans, may be able to settle on top of the flat types.
In his Cook’s tour of the invertebrate world, Buchsbaum includes bryozoans in a chapter entitled “Lesser Lights,” along with rotifers (wheel animals), brachiopods (lamp shells) and certain types of worm. His justification for lumping these disparate groups together is that “they have a small number of species or of individuals; the members are of small size; they constitute no important source of food or of disease for man; and they illustrate no principle of theoretical interest that is not as well shown by other phyla”—all good anthropocentric reasons. Much like the minor prophetic books of the Bible, these groups are relatively little known but have interesting stories to tell.
Buchsbaum’s criteria for relegating bryozoans to the status of invertebrate also-ran might have been acceptable in the 1930s, when he wrote his books, but they no longer apply. The bryozoan star is in the ascendancy, and New Zealanders, especially, have reason to take note.
First, consider the numbers. My colleague the late Alan Horowitz, of Indiana University, attempted to make an inventory of all known bryozoan species in order to track the group’s diversification through time. His tally was 15,000 fossil species and 5700 living ones. Of the living species, 960 (320 still awaiting formal description) occur in the New Zealand region—seven freshwater and the rest marine, the highest regional “bryodiversity” in the world. All these figures are conservative, because new species are being found all the time. When the entire fauna has been described, well beyond my working lifetime, I expect the final count of species living in the New Zealand region will be about 1500.


What is especially remarkable about New Zealand bryozoans is the high concentration of species on some parts of the continental shelf, especially off Spirits Bay, at the northern tip of the North Island, an area that has come to be regarded as a hotspot of marine biodiversity. In 1996, the National Institute of Water and Atmospheric Research (NIWA) conducted a survey of marine life in the scallop-dredging grounds there and in nearby Tom Bowling Bay (see New Zealand Geographic, Issue 46). In shell and rock samples from the survey area, which was no larger than 20 km by 10 km, I was astonished to find 300 species of bryozoan—almost one third of the entire New Zealand bryozoan fauna, and equivalent to the total number of bryozoan species in the Exclusive Economic Zones of Britain and Ireland.
At one Spirits Bay site 140 bryozoan species were found living together. This is the highest number recorded for any seafloor sampling station by NIWA, and represents a diversity that is unmatched anywhere else in the world.
High bryozoan diversity seems to have been a feature of New Zealand’s coastal waters for many millions of years, as attested by the abundance of bryozoan fossils in limestone. Limestone is formed from compressed marine sediments. On parts of the continental shelves of temperate countries such as New Zealand, bryozoan skeletons—from hundreds of different species—are the dominant component of these sediments. It has been estimated that Te Kuiti limestones, including those in the Waitomo area, are 50 to 80 per cent bryozoan remains, and Oamaru stone 70 per cent.
In fact, the very best areas in the country for collecting fossil bryozoans are Oamaru and the Chatham Islands, in quarries and on eroded sea coasts. In such places the collector can be a marine biologist and palaeontologist combined, studying live colonies at low tide and picking out exquisitely preserved fossils from nearby cliffs when the tide is too high for rockpool fossicking.

What about the size of these creatures? Most bryozoans are one to a few centimetres in width (if encrusting) or height (if bushy or erect). A few consist of only a single feeding individual with a few daughter buds, and are small enough to live between sand grains. At the upper end of the scale are species that form coral-like clumps ranging from fist size to a metre across. In fact, whereas in the tropics bryozoans are generally dwarfed by the stony corals—the Scleractinia—in cooler, temperate waters they come into their own, both in size and numbers.
In New Zealand, it is not just the size of some bryozoans that is remarkable but also their extent. Off Abel Tasman National Park there are huge beds of a 15–30 cm-high clump-forming bryozoan known as Tasman Bay coral, Celleporaria agglutinans. A second, more brittle variety, called cornflakes coral, Hippomenella vellicata, forms similar but less robust growths. Like coral reefs in the tropics, these bryozoan micro-reefs attract a variety of other invertebrates and collectively form a habitat in which juveniles of commercial fish species such as tarakihi, snapper and John Dory shelter and feed.
Trawl fishers dragged up chunks of Tasman Bay coral in their nets for decades before realising that damage to the bryozoan beds was affecting fish numbers. In 1980, the Ministry of Agriculture and Fisheries closed two areas to trawling to allow the habitat to regenerate. Even so, the beds have been reduced in area from more than 200 km2 to 55 km2.
But the story doesn’t end there. Both types of coral—Tasman Bay and cornflake—have since been found in fossil deposits near Kahurangi Point, in north-west Nelson, dating back some 20–22 million years. There are clear environmental implications: shallow coastal habitats that may have been in existence for millions of years can be damaged and destroyed by human activities in a matter of decades.
Another habitat-shaping New Zealand species is Cinctipora elegans, a member of a bryozoan family which today is found nowhere else in the world. Ranging from offshore Wanganui to Foveaux Strait and out to the Chathams, Antipodes and Bounty Islands, it features strongly on parts of the continental shelf, where it is associated with an increased diversity of bottom-living species. But in eastern Foveaux Strait it is the principal frame-building species in long reeflike structures on the seafloor.

It seems the twiggy branches of Cinctipora, plus many associated bryozoans and communities of tubeworms and molluscs, trap sand and gravel in their interstices, gradually building up mounds that can be 10 km long, 500 m wide and half a metre high. Living on these mounds is a variety of marine life, including Bluff oysters.
From time to time Bluff oyster stocks are decimated by a protozoan parasite called Bonamia exitiosus, but fishers have noted that oysters on the bryozoan reefs are not affected as severely as those on the seabed. Perhaps the chemical defences given off by the resident fauna of the reefs somehow inhibit the parasite and promote oyster health.
This idea of chemical-related health in the sea is fairly new and has yet to be tested in laboratory conditions. But it is a sobering thought that the very act of dredging for oysters may actually destroy the salubrious character of the habitat in which they occur. NIWA researchers are looking at ways of both monitoring environmental health and mitigating the damage done during oyster harvesting.
[chapter break]
Buchsbaum wrote that bryozoans “constitute no important source of food or of disease for man.” I have to agree with him on the food count. With their calcium carbonate skeletons, bryozoans would be awfully crunchy to eat.
The question of disease is more interesting. There are documented cases of fishers in the North Sea getting contact dermatitis from handling bryozoan by-catch, but this is more in the nature of an allergic reaction than a serious malady.
Rather than causing diseases, bryozoans may, in fact, contain potential cures. A bryozoan-derived biochemical called bryostatin 1 is in the second phase of human clinical trial in North America and Europe for the treatment of several cancers (melanoma, lymphoma, renal cell carcinoma and ovarian and breast cancer). This is the most advanced level of development so far reached by a pharmacological agent from any marine invertebrate, although many others, especially from sponges and sea squirts, are also being considered as anti-cancer drugs.


Bryostatin 1 was first isolated from specimens of the near-cosmopolitan marine-fouling bryozoan Bugula neritina. This species probably hails from the Mediterranean Sea, but has been transported by ships to ports around the world, including New Zealand, where it was first reported in Wellington in 1949. It is now found at most ports from Opua to Dunedin, where it is instantly recognisable from its burgundy-coloured bushy tufts on pilings, moorings, boat hulls and seaweeds, in company with any of 18 other alien bryozoan species. Late spring through summer is the best time to look for Bugula colonies, as they tend to die back in winter.
Intriguingly, Bugula doesn’t produce bryostatin 1 everywhere it occurs in the world, and it is now known there are different genetic populations, each with symbiotic bacteria living in the body cavity of the individual animals of which a colony is composed, making the species distasteful to fish. The chemical is produced by the bacteria, which cannot live apart from their bryozoan hosts.
In recent years, research teams at Canterbury and Waikato Universities have been testing New Zealand bryozoans for other forms of bioactivity. Do they have antibiotic, antifungal, antiviral or anticancer properties? Are they cytotoxic? A dozen species, including the cornflakes bryozoan, have shown promising results, warranting further testing.
“[Bryozoans] illustrate no principle of theoretical interest that is not as well shown by other phyla,” continues Buchsbaum. Recent research would definitely gainsay this assertion. In 1975, a British researcher determined that the retractor muscle which pulls the bryozoan’s feeding crown of tentacles—the lophophore—back into the boxlike body wall can contract at speeds exceeding 20 times its own length per second, making it the fastest-contracting muscle known.
The lophophore tentacles are remarkable in themselves. They are hollow, and either a pair or all of them act as vasa deferentia, conducting sperm from testes in the body cavity and releasing these from a pore at their tip. Thus they act not only as feeding organs and gills, but also as the animal’s reproductive apparatus.
[sidebar-1]
Bryozoans also exhibit an amazing plasticity of structure and function among the individuals that make up a colony. This phenomenon, known as polymorphism (“many forms”), is even more advanced than among the planktonic colonial members of the phylum Cnidaria (which includes siphonophores, the Portuguese man-of-war and the bythe-wind sailor), in which individuals are specialised for feeding, breeding and stinging.
Individuals within a bryozoan colony are called zooids, of which there may be several types (or polymorphs). The main two are autozooids, which are responsible for feeding, and have a lophophore and gut (together called a polypide), and kenozooids, which lack a polypide and contribute only to the structure of the bryozoan colony.
Usually, zooids are hermaphroditic (both male and female), their sperm maturing before their ova. Some bryozoan species have separate female and male zooids that differ in shape and in polypide form from the feeding zooids. Purely reproductive zooids may lack a polypide altogether and have tiny lophophores with reduced numbers of very long and/or very short tentacles whose sole function is sperm release. Such is the case in Odontoporella adpressa, a common New Zealand species that lives almost exclusively on sea-snail shells occupied by hermit crabs. Several species of hermit crab and bryozoan live symbiotically in New Zealand waters, a relationship that seems to have been common for more than 20 million years, since the Miocene period.

In some species, including the pretty purple bryozoan Akatopora circumsaepta, the presence of the hermit induces the encrusting colony to grow beyond the confines of the snail shell, even to the extent of continuing its coiled shape. The bryozoan probably benefits from the arrangement by having access to a ready source of food—the “crumbs” from the crab’s meals—while the crab may benefit by having a permanent “construction crew” on site: the continuing expansion of its home means it doesn’t have to shift premises so often.
Zooids may be modified to function as non-feeding rootlets and stalks for the attachment of bushy colonies, as spines for protection or to control water flow, and possibly as active defenders of the colony. The latter are particularly interesting, as their abundance in some bryozoans raises the question of whether the colony exerts some form of control over their activity.
Charles Darwin was the first to consider the possibility that some zooids fulfil a defence role, the most likely candidates being avicularia. The operculum of these small, modified zooids functions as a mandible, a fan or even a whip. The most striking type of avicularium looks like a bird’s head (hence the name, from avicula, meaning “little bird”). It can rock, and its mandibles can bite and hold organisms which get too close to the colony.
In the encrusting orange bryozoan Beania magellanica, which can be found under low-tidal boulders or on the hold-fasts of kelp, the paired avicularia are quite large. Once, observing a live colony under a microscope, I watched a small copepod crustacean creep across the open gape of an avicularium jaw, triggering its sudden closure. The copepod struggled, and finally succeeded in freeing itself, but left with a dent in its armour.

Amazingly, in spite of the many observations made of avicularia since Darwin’s day, no one has been able to prove a defence role. It is clear that they do not catch food suitable for bryozoans, so perhaps their main task is to discourage larvae of small creatures from settling on the colony. Some avicularia, set near the feeding apparatus, are small enough to suggest they may be taste receptors.
Avicularia in which the operculum is whip-like are known as vibracula. When disturbed, all the vibracular whips in a colony lash in unison, possibly to waft any foreign object away. In some families of free-living bryozoans, these whips give the colony the power of mobility. New Zealand has half a dozen such species, the largest about a centimetre across, the smallest the size of a lentil. Looking like the offspring of a union between a discus and a circular saw, colonies of these species inhabit sandy or muddy-sandy bottoms beyond wave-break. Bristle-like whips, or oars, grow on the domed, upper-colony surface as well as around the edge. If necessary, entire colonies can slowly lurch away from unpleasant situations (top speed is one metre per hour), right themselves if they get overturned and uncover themselves if buried.
Roaming bryozoans pose a question on behalf of all their kind: how can they achieve coordinated nervous control across whole colonies or parts of colonies? It took very careful thin-sectioning of zooids of the cosmopolitan seaweed encruster Electra pilosa (common along New Zealand’s coasts) with diamond knives and examination with an electron microscope for Parisian scientist Geneviève Lutaud to determine that, while bryozoans have no colonial nerves as such, short cells lodged in the pores which connect adjacent zooids form basic synapses with cells of the nerve network (or plexus) contained within each zooid. So while bryozoans are nearly brainless, they do not lack coordination.
Researchers have also observed the transmission of stimuli between zooids. The strength of the colony’s response tends to correlate with the strength of the stimulus. For example, in Australian Selenaria maculata, one of the mobile bryozoans, strong light triggers electrical impulses that cause a coordinated response of the oarlike bristles around the edge of the colony, resulting in the colony being lifted off the bottom and propelled towards the light.
New Zealand also has a large roaming bryozoan, Otionellina squamosa, which is commonly found on clean sandy bottoms in the Hauraki Gulf. This species would certainly lend itself to further study of bryozoan colonial behaviour.


Another intriguing issue, for which bryozoans are proving model study subjects, has to do with how organisms meet their energy costs. Polymorphs are energetically expensive to support because they don’t feed. Several years ago, some Wellington colleagues and I reviewed what was known about bryozoan energetics, and concluded that bryozoans lead an energetically marginal existence, and that there must be considerable fine-tuning in balancing the energy budget. Each bryozoan species is faced with the “choice” of investing in non-feeding polymorphs (for defence or movement), or in extra physical strengthening of the colony by calcium carbonate, or in having more feeding zooids to put nutrient resources into reproduction.
“Resource partitioning” is a hot topic in biological circles right now. Drew Harvell and her students at Cornell University, New York, have been able to measure precisely the amount of energy invested in growth, reproduction and protective devices in the cosmopolitan kelp-encrusting bryozoan Membranipora membranacea. What makes this species an ideal subject is that its defences—chitinous spines on the exposed frontal surfaces of the zooids—are, unusually, inducible. Whenever a particular sea-slug predator is present, the spines are produced: if the predator is absent, more energy goes into growth and reproduction.
The sea slug in question is not found in New Zealand waters, so the local Membranipora, commonly found on Ecklonia kelp, never produces frontal spines.
Bryozoans are also in the forefront of another hot research topic: embryo cloning. A diverse group of animals armadillos, parasitoid wasps and all bryozoans of the order Cyclostomata reproduce by polyembryony, a process in which a single sexually produced embryo splits into multiple embryos, resulting in a batch of genetically identical offspring, or clones. As a reproductive strategy, polyembryony appears to fly in the face of current ideas on the advantages and liabilities of sexual and non-sexual reproduction, hence its very existence is paradoxical. Theories to explain the prevalence of sexual reproduction usually contend that organisms which cannot predict the environment that their offspring will encounter benefit by producing genetically diverse young, which is accomplished by sexual reproduction. With polyembryony, there is no genetic diversity among the offspring. It’s like buying one lottery ticket, then copying it hundreds of times. Common sense suggests it is smarter to have many different lottery tickets, and thereby increase your chances of winning (or, in the lottery of life, surviving).

For parasitoid wasps, which lay their eggs inside caterpillars, there may be benefits in polyembryony. Only some of the clones feed on the caterpillar host; the remainder become protective soldiers, eating other parasitoids that invade the same host. The feeding clones are thus able to monopolise the food supply to complete their development.
The benefit of polyembryony to bryozoans, however, is far from clear, though some suggestions have been made. Few eggs are produced in any given colony, and, since the likelihood of fertilisation by sperm from a distant (or even neighbouring) colony is small, it may be advantageous to multiply, by splitting, the few eggs that are fertilised.
Whatever the explanation, the order Cyclostomata (sole survivor of the bryozoan class Stenolaemata, which had its heyday in the Palaeozoic) is the largest and most ancient group of organisms in which polyembryony occurs. This fact alone argues that attention be given to this group of animals to resolve the paradox of polyembryony. New Zealand has an especially diverse cyclostome fauna, comprising some 100 species, so this would be an ideal part of the world in which to study the phenomenon.
If Ralph Buchsbaum were alive today, I doubt he would be chagrined to learn that bryozoans have overturned all his criteria for minor phylumhood: he would rejoice. As do all of us who love invertebrates and marvel at the myriad ways in which they delight and challenge our imaginations.
Long live MAGS, stimulating science teachers, supportive parents, the memory of Ralph Buchsbaum, and the spirit of scientific inquiry.

Freshwater foulers eutraordinaire
Not all bryozoans live in the sea. At least seven species live in New Zealand’s lakes, ponds and rivers, where their presence goes largely unnoticed. The thin brown threads that make up the colonies are hard to see unless en masse, when they take on the appearance of slippery brown weeds.
Freshwater bryozoans are hard to identify and hard to study, yet in some habitats they are the dominant animals and among the chief foulers of artificial structures.
The story of bryozoan fouling goes back to at least the 19th century, a time when the industrial revolution was transforming Europe. Bryozoans started colonising the complex networks of underground pipes supplying many great European cities with drinking water. The large-bore pipes were perfect homes for them. The pipes’ dark interiors provided the steady currents and hard, stable surfaces required for colony growth, as well as excluding light and major predators. Drinking water was largely untreated at the time, so the water flowing through the pipes teemed with the microscopic river life on which freshwater bryozoans feed.
By the 1880s, bryozoans were creating problems in water pipes throughout much of Europe. The situation in Hamburg, as reported by German biologist Karl Kraepelin, was typical. Like fat-clogged arteries, the city’s water pipes had been diminished in calibre by masses of bryozoan colonies. Water delivery to many homes had been slowed to a trickle—or halted altogether—by the sheer quantity of colonies in the pipes.
Maintenance workers, whose task it was to return pressure to the mains by removing the colonies, named the stuff they were collecting Leitungmoos, “pipe moss,” believing it to be an aberrant, light-shunning plant.
Kraepelin soon discovered that bryozoans weren’t the only creatures lurking in the pipes. He devised a method of removing water samples from anywhere in the pipe system, hoping to find blind crustaceans of the kind normally found in caves. Living among the bryozoans in his samples he found an incredible diversity of life: dozens of species of mollusc, worm, hydroid, leech and sponge. One aquatic species of slater-like crustacean was particularly abundant; Kraepelin recorded it swarming over the bryozoans “in ugly crowds.”
At the top of the food chain were several species of fish. Eels, in particular, were present in their thousands, and would sometimes clog the pipes in a Medusa’s head of writhing forms.
As well as being a major inconvenience, the pipe fauna posed a serious health risk. At the onset of each winter, hundreds of tonnes of bryozoan colonies would simultaneously die off and begin to rot within the pipes, resulting in tap water that was said to be undrinkable even to horses, and which may well have been a source of disease outbreaks. Something had to be done, and it was: the widespread introduction of sand and gravel filtration effectively solved the problem by depriving the bryozoans of food.
Fouling by freshwater bryozoans is by no means of only historical interest. These animals still afflict municipal operations around the world, as well as colonising industrial water intakes and aquaculture facilities. More chillingly, in the 1980s colonies of one species became a significant fouling problem in the vital cooling circuits of a French nuclear power plant.
Closer to home, bryozoans are a continuing source of fouling in Dunedin’s Southern Reservoir, where each year colonies of several species grow in water pipes and water-treatment tanks. During summer, every available surface is transformed into a soft brown carpet of bryozoans, which wave about in the water currents like tussocks on a windy hillside.
Between September and April, council staff are forced to shut down the water supply every six weeks or so and painstakingly water-blast and hand-scrub the surfaces of tanks and pipes to remove the pests. Such maintenance an expensive and irksome disruption to the smooth running of the facility.

The culprits, the freshwater bryozoans Paludicella articulata and Plumatella repens, have been known to be living in Dunedin reservoirs for at least a century. (The first published record was by University of Otago professor Augustus Hamilton, who discovered one of the species when fragments of it came out of his tap.) However, their growth has reached plague proportions in Southern Reservoir only since 1996.
Plumatela is found in most Dunedin reservoirs, and is widespread throughout New Zealand, but it causes fouling problems only in Southern Reservoir. More enigmatic is Paludicela, a northern-hemisphere species whose only known habitat in the entire southern hemisphere is Southern Reservoir.
What is so special about this reservoir that freshwater bryozoans should be so prolific there? Could it be simply that Paludicella arrived from 19th-century Europe along with the machinery and pipes used in the construction of the reservoir, and has survived a century of attempts to get rid of it?
To try to solve the puzzle, a team of Otago bryozoologists have mounted a research programme to examine the freshwater bryozoans of Dunedin’s reservoirs. They have mapped the distribution of the various fouling species, and over the past two years have monitored the bryozoans’ life cycle. Finding out how these bryozoans live may assist in controlling their growth.
But controlling the growth of existing colonies is only half the battle. Both species undergo resting stages, during which they are extremely resistant to environmental stresses. In adverse conditions, they produce seed-like structures, called hibernacula, which can remain dormant for long periods.
The tiny, knobbly hibernacula of Paludicella can survive heating, freezing, drying out, grinding and treatment with acid. Their very robustness gives weight to the argument that the species could have been introduced to the country as stowaways with the reservoir materials and construction tools.
The research group has tested the effectiveness of ultraviolet light and ultrasound in controlling bryozoan growth, and is shortly to investigate the effect that chemicals used in the council’s recently adopted membrane filtration treatment process have on both adults and hibernacula.
The Dunedin City Council Water and Waste Business Unit hopes that a safe treatment will be found to limit or actually stop settlement and growth of freshwater bryozoan colonies at Southern Reservoir. In the process, scientists hope to find out more about these unusual, understudied, but fascinating organisms.